


Coming up for air
Within a few hours of the naval sonar drill, reports arrived of stranded beaked whales appearing over many kilometres along the coast. These animals showed signs of decompression sickness, also known as ‘the bends’.
Post-mortems on these animals revealed gas and fat bubbles in their bones and tissues.
The deeper you dive, the more the pressure forces nitrogen and oxygen from your lungs to dissolve into your body tissues. If you then surface too quickly, these gases can come out of solution and form bubbles in your blood. These can block smaller blood capillaries, cutting off the oxygen supply to the affected tissues. Decompression sickness is a recurrent risk amongst scuba-divers who breathe compressed air, and breath-holding ‘free-divers’ who make too many consecutive dives.

We have a diving reflex like other mammals. As the water hits our face, our heart slows and muscles under the skin contract, shunting blood into the centre of our body. Water pressure increases by 1 Atmosphere for every... more 10m depth. At 2 Atmospheres, the air in our lungs is half its original volume. By 50 metres (5 Atmospheres), gaseous oxygen and nitrogen dissolves into our body tissues, and fluid floods into our lungs. The human free-diving depth record is 214 metres (Image: Wikimedia Commons)
In contrast, beaked whales routinely hunt for an hour below 1000m, using echolocation. These ‘extreme divers’ do not normally experience decompression sickness, although fossils from early in their evolutionary history show that they were not immune to these problems. X-rays of the fossilised bones of more primitive whales show regions where bubbles formed inside a capillary, damaging the bone tissue and leaving a tell-tale signature.
Whale embryos initially develop rear limb buds, like land mammals. These structures are reabsorbed back into the body later in development. The fossil record, along with DNA studies, reveal that whales’ closest living relatives are cows and hippos, which share their same four-legged (tetrapod), hoofed, land-dwelling ancestors.
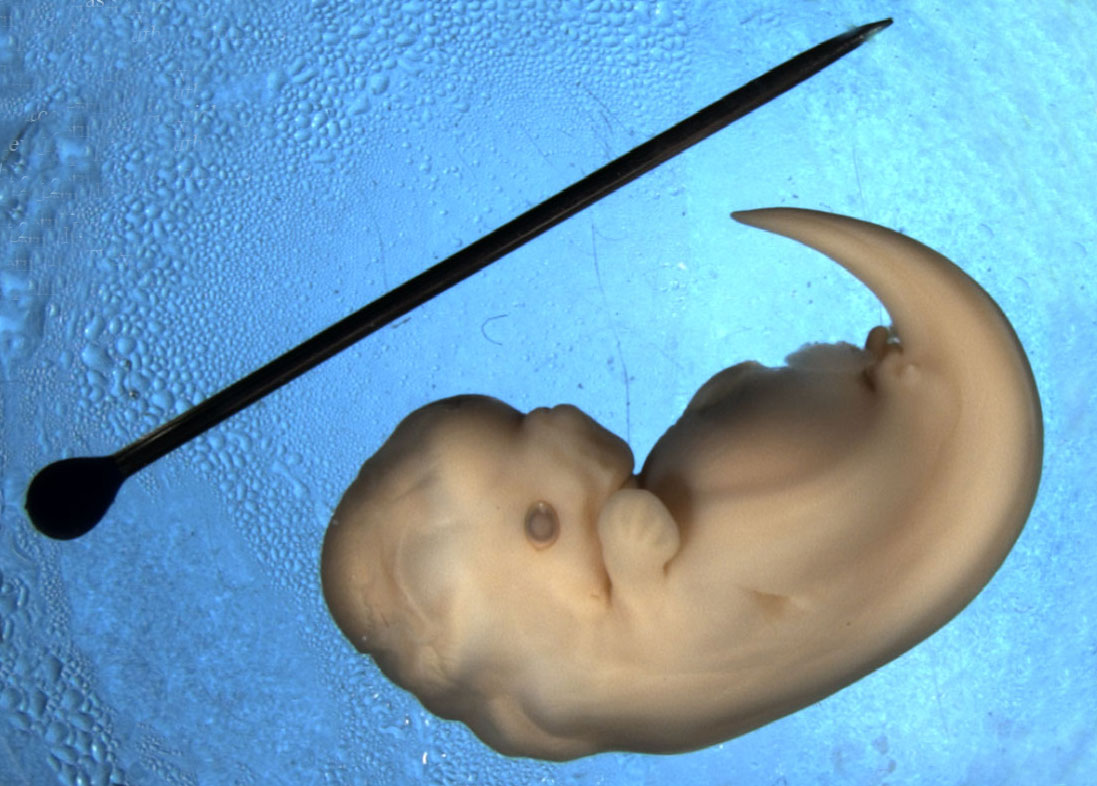
The hind limbs of this Spotted Dolphin embryo (Stenella frontalis) are visible as small bumps (limb buds) near the base of the tail. (Image: Wikimedia Commons)
This raises some puzzling questions:
– Why did whales’ ancestors take to the water after 300 million years on land?
– Why didn’t they re-evolve gills?
– How can they dive for so long without getting ‘the bends’?
Why did whales’ air breathing ancestors take to the water?

These North Ronaldsay sheep are descended from an Orkney population farmed here since Neolithic times. They graze along the shoreline, feeding almost exclusively on seaweed. Their rumen stomachs have an adapted bacteria... morel population which enables them to digest marine algae (Image: Wikimedia Commons)
The land-dwelling ancestors of whales may have first waded into the sea to escape from predators on land. Shallow coastal areas offered a relatively safe haven with little competition for the new food resources available in or near the water. This initial stage would have enabled these semi-aquatic ancestors of modern whales to adapt their digestive systems to a marine food source.
Fossils from the early Eocene (52Ma) show a succession of increasingly aquatic forms. From crocodile-like and otter -like amphibious hunters, developmental changes remodelled their breathing, senses, kidney function and limbs to survive better in water. By 40Ma, these early whales had flippers, a fluked tail, and could mate, birth and suckle their young without leaving the water.
At the Eocene-Oligocene boundary (around 36Ma), movement of the continental plates opened up the deep waters of the circum-Antarctic ocean. This offered new ecological roles for the deeper-diving whales. Many new whale species appeared, including ancestors of the filter-feeding baleen whales and toothed whales that hunt in deep waters using echolocation.
Why didn’t whales re-evolve gills?

A sperm whale (Physeter macrocephalus) begins a dive; Gulf of Mexico. Adaptations for cold, deep waters include insulating blubber, lungs designed to collapse under pressure, and locomotion. The fluked tail is a super-e... morefficient ‘caudal oscillator’; both the up and down strokes generate lift, like a birds’ wing. These and other whale and seal species dive deep both to forage and to escape from killer whale (Orcinus orca) attacks (Image: Wikimedia Commons)
The ability to breathe underwater like fish seems at first like a requirement for life in the sea. However despite their lack of gills, whales and dolphins are highly effective predators in both shallow and deep water.
Modern whales’ warm bodies enable their fast reflexes for hunting. Whilst swordfish and tuna have some warm muscles, most of their tissues are at sea water temperature. Were their whole bodies warm, the heat loss from their gills would be energetically too costly.
Fish gills develop from the ‘branchial arches’; bulging structures in the early vertebrate embryo. These same tissue bulges give rise to the lower jaw, the middle ear, hyoid bone and larynx in the throat of humans and other mammals. For whales and other mammals to form gills would require that they develop new embryonic structures; this would render redundant the lungs with their vast area of vascular tissue.
Breathing air enables whales to use vocal signals to coordinate their social groups and attract mates. Like land mammals, the baleen whales make vocal calls by passing a controlled air flow through the larynx. Echolocation, the alternative means of producing sound used by dolphins and other toothed whales, also requires air. Their ‘sonic lips’ generate calls in an air-filled nasal passage. Whilst many fish make sounds, their vocal abilities are simple and limited.
How do they dive for so long without getting ‘the bends’?
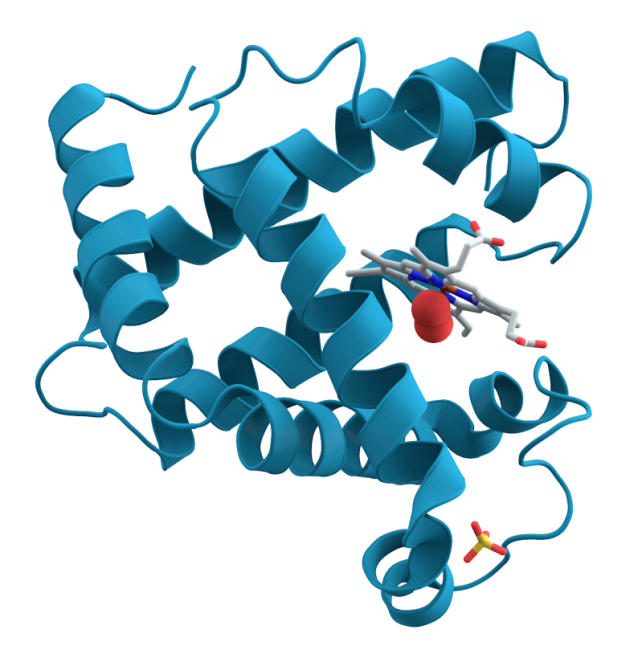
This diagram shows how myoglobin forms ‘alpha-helical’ spirals around a ‘haem’ co-factor. Haem’s ring-structure holds an iron atom, carrying an electrostatic charge. This attracts and holds an oxygen molecule ... more(red spheres). As carbon dioxide builds up it dissolves to form carbonic acid. This change of pH, alters the electrostatic balance, prompting myoglobin to release its oxygen. The myoglobin protein’s high positive charge also steadies the pH when cells break down sugars without oxygen and produce lactic acid (Image: Wikimedia Commons)
All mammals store oxygen in their muscles using a protein called myoglobin. Sustained activity during long foraging dives requires a lot of oxygen. Deep divers have much higher muscle myoglobin concentrations than land mammals, giving them substantial oxygen reserves.
Modern diving mammals, and deep diving fish such as tuna, have also modified their myoglobin. As early whales began to explore the deeper waters, selection resulted in better survival from individuals whose myoglobin carried a stronger positive electrostatic charge. Like positive magnetic poles, these ‘supercharged’ molecules repel each other. This keeps them in solution, allowing them to function at high tissue concentrations where most other proteins would clump together.
A supercharged form and high concentration of myoglobin makes it possible for deep diving mammals to return to the surface slowly after a prolonged dive. This behaviour avoids decompression sickness.
However when beaked whales and other species encounter naval sonar at depth, this causes them to ‘panic’ and surface too quickly, inducing ‘the bends’.
Text copyright © 2015 Mags Leighton. All rights reserved.
References
Balasse M et al. (2006) ‘Stable isotope evidence (δ13C, δ18O) for winter feeding on seaweed by Neolithic sheep of Scotland’ Journal of Zoology 270(1); 170-176
Beatty B L & Rothschild B M (2008) ‘Decompression syndrome and the evolution of deep diving physiology in the Cetacea’ Naturwissenchaft 95;793-801
Costa D P (2007) Diving physiology of marine vertebrates’ Encyclopedia of life sciences doi:10.1002/9780470015902.a0004230
Ferguson S H et al. (2012) ‘Prey items and predation behavior of killer whales (Orcinus orca) in Nunavut, Canada based on Inuit hunter interviews’ Aquatic Biosystems 8 (3); http://www.aquaticbiosystems.org/content/8/1/3
Gatesy J et al. (2013) ‘A phylogenetic blueprint for a modern whale’ Molecular Phylogenetics and Evolution’ 66:479-506
Mirceta S et al. (2013) ‘Evolution of mammalian diving capacity traced by myoglobin net surface charge’ Science 340;1234192
Nery M F et al. (2013) ‘Accelerated evolutionary rate of the myoglobin gene in long-diving whales’ Journal of Molecular Evolution 76;380-387
Noren S R et al. (2012) ‘Changes in partial pressures of respiratory gases during submerged voluntary breath hold across odontocetes; is body mass important?’ Journal of Comparative Physiology B 182;299-309
Orpin C G et al. (1985) ‘The rumen microbiology of seaweed digestion in Orkney sheep’ Journal of Microbiology 58(6); 585-596
Rothschild B M et al (2012) ‘Adaptations for marine habitat and the effect of Jurassic and Triassic predator pressure on development of decompression syndrome in ichthyosaurs’ Naturwissenchaften 99;443-448
Steeman M E et al. (2009) ‘Radiation of extant cetaceans driven by restructuring the oceans’ systematic biology 58;573-585
Thewissen J G M et al (2007) ‘Whales originated from aquatic artiodactyls in the eocene epoch of India’ Nature 450;1190-1195
Thewissen J G M et al (2006) ‘Developmental basis for hind-limb loss in dolphins and origin of the cetacean body plan’ Proceedings of the National Academy of Sciences USA 103(22); 8414–8418
Thewissen J G M and Sunil B (2001) ‘Whale origins as a poster child for macroevolution’ Bioscience 51(12);1037-1049
Tyack P L (2006) ‘Extreme diving of beaked whales’ Journal of Experimental Biology 209;4238-4253 Naturwissenchaften 99:443-448
Uhen M D (2010) ‘The origin(s) of whales’ Annual Review of Earth and Planetary Sciences 38;189-219
Uhen M D (2007) ‘Evolution of marine mammals; back to the sea after 300 million years’ The Anatomical Record 290;514-522
Williams T M (1999) ‘The evolution of cost efficient swimming in marine mammals; limits to energetic potimization’ Philosohical Transactions of the Royal Society of London series B 354;193-201
The reptile that almost became a fish
‘It’s head is as long as I am tall!’
‘What is it?’
‘Hmm… A giant fish? A lizard? Don’t know.’
‘Well I know! It’s a sea dragon!’
It is spring, 1811, the morning after a storm. Mary is 12 years old, and fearless. She edges across the cliff to where her brother is already working to free the fossil bones. Fragments of weathered mudstone clatter down onto the beach. The eye sockets of the huge skull are wider than the span of her hand.

A 185 million year old fossil of Ichthyosaurus acutirostris beside ammonites (Harpoceras falcifer). This specimen shows the distinctive downward (hypoocercal) bend of the spine into the lower tail fluke, characteristic ... moreof this reptile group. The outlines of the fluked tail and dorsal fin are visible; these were supported by cartilage rather than bone as in modern fish. The huge eye sockets (relative to its body size) enabled these animals to hunt by sight for shellfish, small fish and squid in dimly lit or murky waters (Image: Wikimedia Commons)
During the 200 million years that dinosaurs roamed the land, the oceans were ruled by formerly land-dwelling reptiles. Of these, ichthyosaurs adopted dolphin-like forms, plesiosaurs became sea lion-like, and mosasaurs occupied crocodile-like ‘ambush’ predator roles.
Of these, ichthyosaurs were arguably the most successful. Their multiple adaptations to a fully aquatic life included huge eyes, a stiffened, fluked tail, and the ability to mate and give birth to live young in water. Much as killer whales do today, these predators structured the marine ecosystem. Many came to resemble the modern whales, dolphins and tuna fish that now fulfil similar ecological roles. The convergence of body forms between some ichthyosaur species and tuna are particularly astonishing because these reptiles were air breathers.
Why did icthyosaurs evolve to look like modern marine animals?

Ichthyosaurs first colonised the sea 250Ma ago. The earliest known aquatic ichthyosaur, the otter-like Utatsusaurus hataii (top) fed on fish and shellfish in shallow water. After the Cretaceous-Tertiary mass extinction ... more(65Ma), the land-based ancestors of modern whales also took to water. The early whale Kutchicetus minimus (bottom) had an otter-like ecological role, and converged to evolve a similar body form. Both had an undulating swimming style which was in the horizontal plane (like an eel) for Utatsusarus, and vertical for Kutchicetus. This reflects the style of locomotion inherited from their respective ancestors (Images: Wikimedia Commons)
Life in water poses a specific set of challenges. These marine reptiles fulfilled similar ecological roles to whales, and in time evolved body forms similar to these modern mammals. This is known as convergence.
Like whales and tuna, ichthyosaurs were adapted for long distance energy-efficient swimming. The respective horizontal and vertical strokes of both ichthyosaur and dolphin tails give an equally powerful ‘lift’ in both directions, propelling these animals forward in a near straight line.
A further example of this convergence is seen in the modern otter. Unlike the Ichthyosaurs, both whales and otters had land-based mammalian ancestors with a vertically moving spine, giving them their bounding gait. The whale-like ichthyosaurs moved their tail flukes horizontally, like a modern lizard.
Like whales and tuna, ichthyosaurs were adapted for long distance energy-efficient swimming. The respective horizontal and vertical strokes of both ichthyosaur and , propelling these animals forward in a near straight line.
What can modern animals tell us about ichthyosaurs?

The ichthyosaur Stenopterygius quadriscissus (above), became widespread, in the late Jurassic and early Cretaceous (160-100Ma). Its body shape is similar to that of the Bluefin tuna (Thunnus thynnus (below). Tuna hunt f... moreish and squid at around 500m depth. This is possible because they have a high oxygen intake, fast metabolic rate, warm muscles, an energy efficient swimming style, and a higher heart rate and blood pressure than other fish (Images: Wikimedia Commons)
Modern tuna and lamnid sharks have converged into a similar ‘deep water sprint-predator’ ecological role. These long distance migrants move constantly at a moderate speed, except for short fast bursts when chasing prey. Chunky vertebrae stiffen their bodies at their core, reducing sideways movements except at the narrow ‘hinge’ before the tail. The continuously active red ‘cruising’ muscles either side of the spine are warm, in marked contrast to most other fish. In combination with large tendons, these muscles work like pulleys, flicking the fluked tail from side to side. The surrounding white muscles give extra power during short ‘sprints’.
The ichthyosaur Stenopterygius was tuna-shaped with chunky stacked vertebrae. This stiffened body form suggests that these reptiles converged on the cruise-and-sprint deep water hunting role of modern tuna and lamnids.
Did ichthyosaurs have warm muscles like whales and tuna?

Cast of a skeleton of Hawkins’ plesiosaur (Thalassiodracon hawkinsi) from the Lower Lias strata at Street in Somerset; part of England’s Jurassic coast. These rocks are rich in marine fossils of all kinds including ... morefish, ammonites and belemnites (Image: Wikimedia Commons)
The isotopic proportion between the heavy 18O and the light 16O oxygen (the ratio is given as d18O) in the bones of living fish and marine animals decreases as body temperature increases. In principle we can use cold-blooded fish fossils as a ‘thermometer’ to indicate the water temperature, and compare this against isotope-predicted body temperatures for other fossil animals from the same rocks.
Oxygen isotope data allows us to infer that Jurassic plesiosaurs and ichthyosaurs had body temperatures of around 35⁰C, much higher than that of their environment. This indicates that they could generate heat, and may well regulated their body temperatures independently of their environment (homeothermy). Modern warm bodied marine animals have to conserve their body heat. This means it is reasonable to infer that ichthyosaurs used similar methods such as a counter-current blood circulation system and/or heat-insulating blubber.
What happened to the ichthyosaurs?

Plotosaurus bennisoni; a mosasaur from the Upper Cretaceous of North America. Most mososaurs lived in shallow coastal waters, although after the disappearance of the ichthyosaurs, some evolved into similar deep water sp... morerint predators. Plotosaurus had crescent-shaped tail flukes, equipping this animal to whale-like fast pursuit behaviour (Image: Wikimedia Commons)
Ichthyosaurs dominated the world’s oceans for around 150 million years, but then disappeared from the fossil record after the mid-Cretaceous (around 95Ma). The cause of their sudden extinction remains a mystery. The empty ecological roles that this created were later filled by mosasaurs; relatives of modern monitor lizards including the Komodo dragon. In turn these reptiles died out during the Cretaceous-Tertiary mass extinction (65Ma), making way for the later evolution of modern whales, dolphins and tuna.
Text copyright © 2015 Mags Leighton. All rights reserved.
References
Benson, R.B.J. and Butler, R.J. (2011) Uncovering the diversification history of marine tetrapods: ecology influences the effect of geological sampling biases In Comparing the Geological and Fossil Records: Implications for Biodiversity Studies ( A.J. McGowan and A.B.Smith, eds). Geological Society of London Special Publications 358, 191-208.
Bernal, D. et al. (2005) Mammal-like muscles power swimming in a cold-water shark. Nature 437: 1349-1352.
Brill, R.W. (1996) Selective advantages conferred by the high performance physiology of tunas, billfishes and dolphin fish. Comparative Biochemistry and Physiology, A 113, 3-15.
Brill, R.W. et al.(2005) Bigeye tuna (Thunnus obesus) behavior and physiology and their relevance to stock assessments and fishery biology. Collective Volume of Scientific Papers ICCAT 57, 142-161. Online;http://www.iccat.es/en/pubs_CVSP.htm
Dickson, K.A. and Graham, J.B. (2004) Evolution and consequences of endothermy in fishes. Physiological and Biochemical Zoology 77, 998-1018.
Donley, J.M. (2004) Convergent evolution in mechanical design of lamnid sharks and tuna. Nature 429, 61-65.
Fröbisch, N.B. et al. (2013) Macropredatory ichthyosaur from the middle Triassic and the origin of modern trophic networks. Proceedings of the National Academy of Sciences, USA 110, 1393-1397.
Graham, J.B. and Dickson, K.A. (2004) Tuna comparative physiology. Journal of Experimental Biology 297, 4015-4024.
Houssaye, A. (2013) Bone histology of aquatic reptiles: what does it tell us about secondary adaptation to an aquatic life? Biological Journal of the Linnean Society 108, 3-21.
Korsmeyer, K.E. et al. (1996) The aerobic capacity of tunas; adaptation for multiple metabolic demands. Comparative Biochemistry and Physiology, A 113, 17-24.
Lindgren, J. et al. (2007) A fishy mosasaur: the axial skeleton of Plotosaurus (Reptilia, Squamata) reassessed. Lethaia 40,153–160.
Lindgren, J. et al. (2011) Landlubbers to leviathans: evolution of swimming in mosasaurine mosasaurs. Paleobiology 37, 445-469.
Lingham-Soliar, T. and Plodowski, G. (2007) Taphonomic evidence for high-speed adapted fins in thunniform ichthyosaurs. Naturwissenschaften 94, 65-70.
Malte, H. et al. (2007) Differential heating and cooling rates in bigeye tuna (Thunnus obesus Lowe); a model of non-steady heat exchange. Journal of Experimental Biology 210, 2618-2626.
Motani, R. et al. (1998) Ichthyosaurian relationships illuminated by new primitive skeletons from Japan. Nature 393, 255-257.
Motani, R. (2002) Scaling effects in caudal fin propulsion and the speed of ichthyosaurs. Nature 415, 309-312.
Motani, R. (2002) Swimming speed estimation of extinct marine reptiles: energetic approach revisited. Palaeobiology 28, 251-262.
Motani, R. (2010) Warm-blooded “Sea Dragons”? Science 328, 1361-1362.
Rothschild, B. M. et al. (2012) Adaptations for marine habitat and the effect of Triassic and Jurassic predator pressure on development of decompression syndrome in ichthyosaurs. Naturwissenschaften 99, 443-448.
Syme, D.A. and Shadwick, R.E. (2011) Red muscle function in stiff bodied swimmers: there and almost back again. Philosophical Transactions of the Royal Society, B 366, 1507-1515.
Thewissen, J.G.M. and Bajpai, S. (2009) New skeletal material of Andrewsiphius and Kutchicetus, two Eocene cetaceans from India. Journal of Paleontology 83, 635-663.